Origin and evolution of biospheres (C4)#
Module leader: Emily Mitchell (Zoology)
Learning objectives:
Understand the basics of aqueous and organic chemistry
Understand the chemistry of atmospheres
Understand chemistry in astrophysical contexts
Be familiar with how the chemistry of environments can be used to detect life
This first introductory lecture will introduce us to how we consider life and evolution in terms of the major evolutionary transitions.
Introduction to biospheres#
How we define life is something that is much debated, and NASA define life as:
Nasa’s definition of life
A self-sustaining chemical system capable of Darwinian evolution
By this they mean that living systems contain molecules that undergo directed chemical transformations (metabolisms) performed by enzyme catalysts. These molecular structures are inherited such that information is transferred through the generations and the best (fittest) survive.
In terms of what constitutes life on this planet currently, the majority of life in terms of biomass terrestrial and plants. There is more deep subsurface biomass (bacteria and archaea) that all the life in the oceans than in the marine realm. Animals consistent a relatively small proportion of life on Earth, and within these its mostly invertebrates such as insects. So when we think about life, we need to keep in mind that most of life both now, and through deep time is not animals, let alone humans that dominate, and for most of Earth’s history it was just microbial.
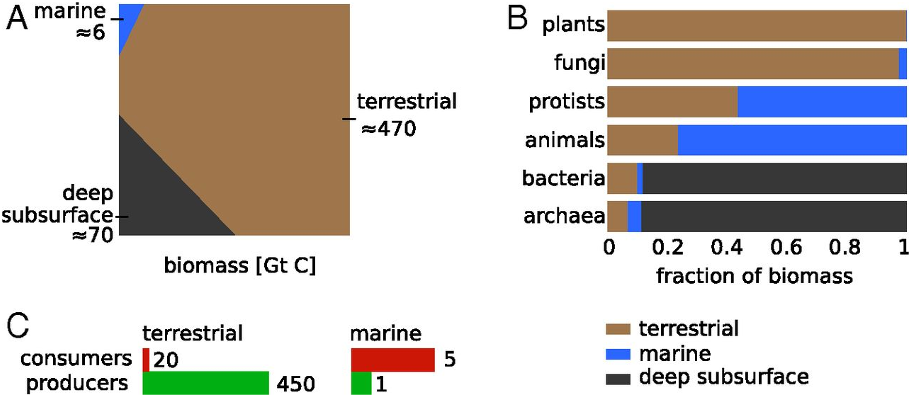
Fig. 1.27 A distribution of terrestrial biomass by its spatial distribution Bar-on+2018.#
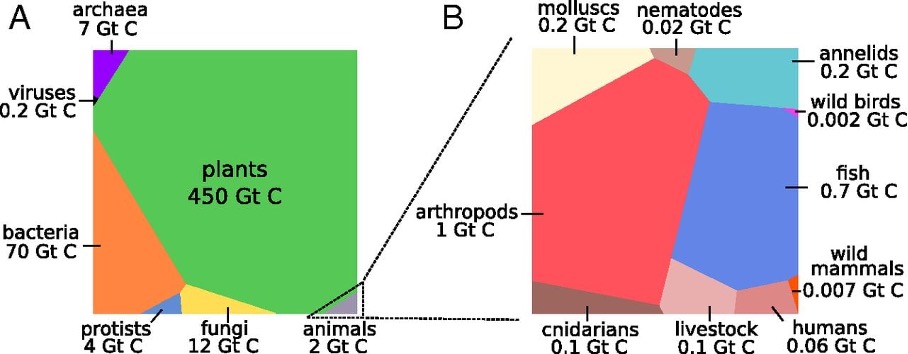
Fig. 1.28 A distribution of terrestrial biomass by type Bar-on+2018.#
Major evolutionary transitions#
In Astrobiology we commonly consider the Drake equations, with the last four focussed on the origins of life and biology.
\(f_l\), the fraction of planets that could support life that actually develop life at some point.
\(f_i\), the fraction of planets with life that go on to develop intelligent life (civilizations).
\(f_c\), the fraction of civilizations that develop a technology that releases detectable signs of their existence into space.
\(L\), the length of time for which such civilizations release detectable signals into space.
However, there are a huge amount of assumptions and biology missing within these: a single term \(f_i\), that is the fraction of planets with life that go onto develop intelligent life encapsulates pretty much all of evolutionary biology, from LUCA to us, yet so there so much about the evolutionary history of life on Earth that we don’t understand or know that more than a single probability is needed. Instead we are going to approach these questions from an evolutionary biological perspective and consider the major evolutionary transitions, because that provides a framework for then going on to think about the probability of such events occurring, and what drives such changes.
Maynard Smith and Szathmary first proposed the seven Major Evolutionary transitions. All of these transitions have common factors, in that for each transition you go from smaller to larger entities, both in terms of physical space, but also in terms of complexity. The smaller, previous steps are often a differentiated part of the larger/later step, and with each step there can be an associated increase in information transmission.
Chromosomes
DNA as genes; proteins as enzymes
Eukaryotes
Evolution of sex
Evolution of multicellularity
Evolution of eusociality
Sociocultural evolution
Abiogenesis#
The processes whereby abiogenesis, that is the transition from abiotic to biotic molecules , occurs is very much debated. In terms of life today, DNA encodes the information through the generations, and replicates using enzymes and proteins such as RNA. This creates a bit of a cycle - if both metabolism, (i.e. the autocatalytic network of reactions needed for replication) and the DNA with the genes are needed for this cycle which came first? Well there are two main ideas. RNA first, which is sometimes called top down and metabolism, or bottom up. Both start with a soup of small carbon-based molecules. In an RNA/gene first model the nucleic acid monomers form then combine. These Polymers then catalyze the formation of more monomers in a feedback leading to the direct assembly of identical polymers. Selection is then thought to promote efficient and the acquisition of additional functions.
In a metabolism/protein first model From the carbon-based molecular soup non-enzymatic catalysts expand the chemical inventory to create networks of reactions. Cyclic reaction sequences provide a pathway to lower energy levels for high-energy electrons that are initially present. Selection leads to increased catalytic versatility and so metabolism continues to evolve to present form. Opponents of RNA first would say that 1) RNA is too complex a molecule to have arisen prebiotically; (ii) RNA is inherently unstable; (iii) catalysis is a relatively rare property of long RNA sequences only; and (iv) the catalytic repertoire of RNA is too limited. Criticisms of metabolism first is the networks need self-replicating abilities with a high degree of fidelity, to ensure that the chemical networks with greater fitness in early Earth survive.
Prokaryote to Eukaryotic transition#
The next major evolution transition is that of biological complexity – the transition from simple cells that lack internal structures, namely prokaryotes such as bacteria and archaea to complex cells with internal structures such as nucleus and other organelles such a mitochondria and in plants chloroplasts. We think that eukaryotes evolved from an endosymbiotic event because:
Mitochondria and chloroplasts are approximately the same size as prokaryotic cells, inside the much larger eukaryotic cells.
The membrane composition of mitochondria and chloroplasts is more similar in composition to prokaryotic membranes than to eukaryotic membranes.
All the genes to produce the mitochondria or chloroplasts are found inside these organelles and are not present eukaryotic nuclear genome.
The mitochondria/chloroplast DNA are circular chromosome like typical prokaryotic genomes, with similar prokaryotic genes, and are not linear like the eukaryotic nuclear chromosomes.
Mitochondria and chloroplasts reproduce by binary fission (the cell splits) whereas eukaryotes cells reproduce by mitosis whereby the cells duplicate before splitting.
Asexual to sexual reproduction#
The evolution of sexual reproduction goes hand in hand with the evolution of eukaryotes and fundamentally changed the nature of evolution. This fundamental change is because sexual reproduction, where the genetic information from two individuals is combined, enables much higher genetic variation, so the rate in which you produce novel genotypes is much higher. These genetic novelties means that sexual produced populations are much better at adapting to changing conditions, such as different environments, or new predators or competitors for the same food source. Sexual reproduction also means that if mutation has a negative effect on fitness then it will get removed from the population relatively quickly.
However, the cost of sexual reproduction is very high, and with mammals such as humans where only half the population produces offspring it is referred to as the two-fold cost of sex. Any sexual reproduction is energetically costly because the organism needs to expand energy to make the gametes and then ensure they get combined with other gametes. The relative costs of sexual reproduction are managed by many Eukaryotic clades by being able to reproduce both sexually and asexually, which includes trees, fungi, invertebrates and vertebrate animals.
Multicellularity#
Multicellularity is any organism that consists of more than one cell, which can be the same or different cells. As such, the evolution of multicellularity isn’t a single transition, because it has occurred independently many times across eukaryotes and bacteria, but not archaea.
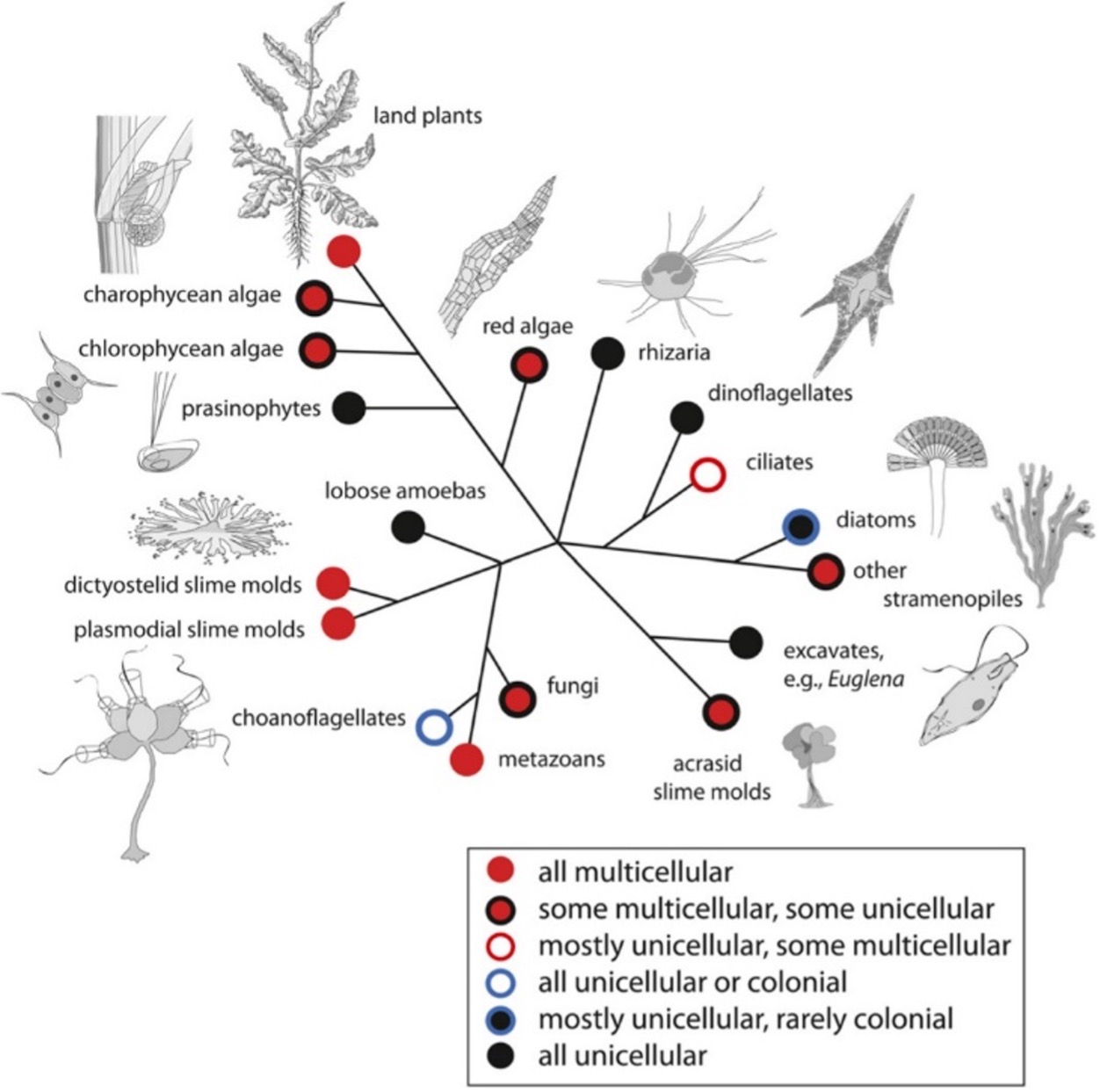
Fig. 1.29 The origins of multicellularity as a simplified phylogenetic diagram for the major eukaryotic clades Niklas, 2014.#
Assessment of Maynard-Smith Major Evolutionary transitions#
How useful are these Maynard-Smith Major Evolutionary transitions when thinking about life elsewhere? There are some fundamental problems with these MET. The first being there is an implication of equivalence, which is often seen in astrobiology papers which treat them all as if they are functionally equivalent, which isn’t correct. For example, some are coupled, such as with the evolution of eukaryotes and of sex, the order of the first two are debated, and all three of the last ones have happened multiple times. Yet, if an organism evolves independently, over and over again, then it is far more likely, so not the same as a transition that has only happened once. There are also key steps that are missing, such as the evolution of oxygenic photosynthesis. So instead, we are going to think in terms of the 3 evolutionary transitions that led to a step-change in terms of the biosphere – that is this the innovations of life that changed the entire planet forever.
Great ecological transitions#
The four Great Ecological transitions are the origins of life, the evolution of oxygenic photosynthesis, the evolution of animals and of land plants.
Origins of life: Lectures 2-6 In terms of where life started, it is very much debated, with submarine and hot springs the most supported ideas. In terms of the steps they can be grouped as:
Catalysis: Simple organic molecules were synthesized from primordial inorganic molecules
Self-Assembly: More complex polymers were constructed from these simple organic molecules
Self-Replication: Certain polymers formed the capacity to be duplicated (enabling inheritance)
Compartmentalization: These molecules became packaged into membranes with unique internal chemistry
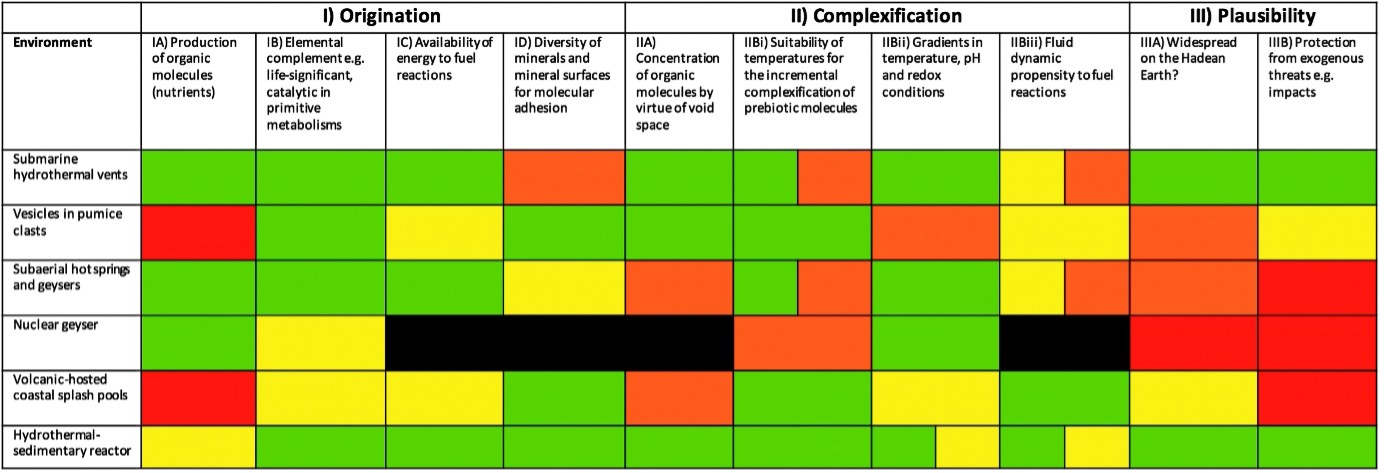
Fig. 1.30 The potentials of the proposed environments for the origin of life inidicated by colours, green is good, red is bad Westall+2018.#
All the different environments have the ability to concentrate molecules, and have either temperature, pH and/or chemical gradients to maintain differences outside equilibriums.
Oxygenic photosynthesis: Lecture 7-8 Photosynthesis has evolved up to 16 different times, including in prokaryotes, yet oxygenic photosynthesis has only evolved once. The introduction of these oxygen producing cyanobacteria led to the build-up on oxygen in the atmosphere around 2.5 billion years ago, in what is known as the great oxygenation event.
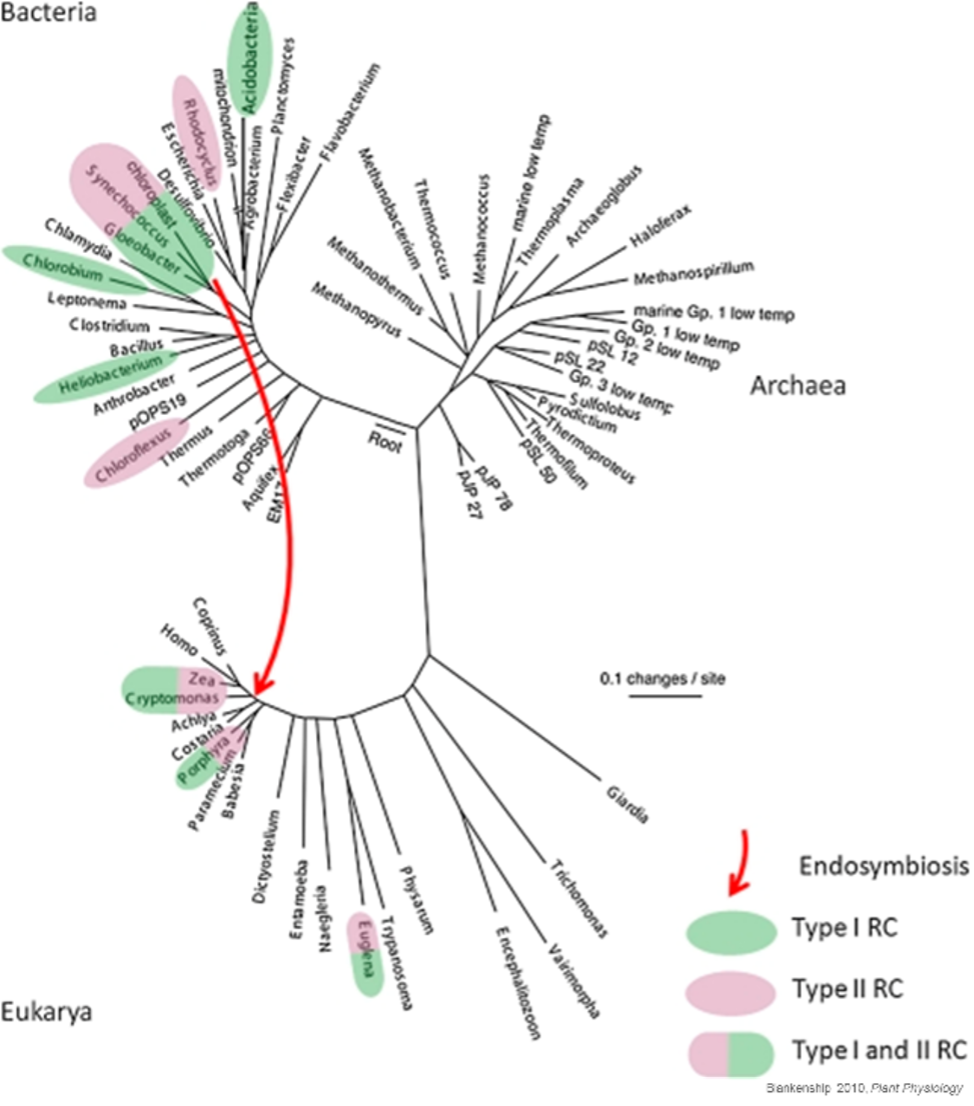
Fig. 1.31 An rRNA evolutionary tree of life, with taxa containing photosynthetic representatives highlighted. The red arrow indicates the endosymbiotic event that formed eukaryotic chloroplasts Blakenship, 2010.#
Animals: Lecture 9 It is only a little under 600 million years ago that animals evolved during the Ediacaran time period. The really big innovation of animals were their ability to move around, and so change the environment around them. The evolution of animals changed the physical environment through their movement, mixing the ocean waters and the seafloor.
Land Plants: Lecture 10 The final really big biosphere change was the evolution of land plants, the biggest one being through oxygenation of the atmosphere, but also through the stabilizing the land masses with their roots, increases in the amount of carbon buried within sedimentary rocks, changes the continental weathering regime, hydrological cycle. Although note that land plants contribute to 50% of the atmospheric oxygen, with marine phototrophs contributing the rest.
Our final two lectures are consider how evolution works in terms of diversity patterns through geological time scales, how things change, what drives these difference and why they change. Part of the difficulty with our current n=1 life is that it’s so hard to tease apart correlation from causation, and so we need to think about the different aspects that impact evolution, why they occur and before we start to consider what evolution will look like on other planets and bodies. In our final lecture we will consider the different mechanisms for evolution in terms of null models, environmental drivers and biotic drivers, before thinking about how these and the interplay between them may vary elsewhere
Key References#
Blankenship, R.E., 2010. Early evolution of photosynthesis. Plant physiology, 154(2), pp.434-438.
Social-cultural evolution#
The final of Maynard-Smiths major evolutionary transitions is the evolution of social culture which is the passing down of learned behaviours, traditions and beliefs within a group. Culture animals include Great apes, especially chimpanzees, cetaceans (Whales, dolphins, porpoises) as well as meerkats and naked mole rats. There is broadly wide agreement that great and some lesser apes have cultures, alongside Cetaceans, but because there are differences to how exactly you define culture, versus learning, people would also argue that birds, rats and most controversially some fish also exhibit culture. And as such there isn’t an absolute number of times that species have evolved their own culture, but it has happened many times.